FW SCOTT LAB Group
Team Leader

Fraser Scott
Emeritus Scientist, Inflammation and Chronic DiseaseWhat We Do
T1D is an autoimmune disease in which the patient's immune system destroys the insulin-producing β-cells in the pancreatic islets. The rise in incidence over the past 50-70 years is not explained by changes in genetic risk, but is thought to be due to factors in the environment including viruses, dietary antigens and gut microbes. All of these agents first encounter the immune system in the gastrointestinal tract. For several decades before retiring in 2019, the aim of my research was to better understand how these factors influence the gut immune system, pancreas inflammation and β-cell health.
Research Activities
Note: Dr. Scott retired in 2019 and the laboratory is closed. The following summarizes some of the major findings and overall conclusions of his group's research.
Environmental control of gut immunity, β-cell health and type 1 autoimmune diabetes (T1D)
During my research career my group was interested in how environmental factors, particularly dietary antigens, influence the development of T1D [1,2]. A major focus was to understand the diabetes-promoting interactions between dietary antigens, the gut immune system and endocrine pancreas by (i) characterizing pathways in gut and pancreas that are abnormal [3-5] and (ii) defining abnormal gut immune responses to dietary and microbial antigens that promote or inhibit diabetes in animals and patients [6,7]. T1D is an autoimmune disease in which the patient's immune system destroys the insulin-producing β-cells in the pancreatic islets. The remarkable ~2-4 fold increase in incidence over the past 60-70 years is not explained by changes in genetic risk, but is thought to be due to factors in the environment. The prime suspects remain enteroviruses, dietary antigens and gut microbes. All of these agents have their first major encounter with the immune system in the gut. Thus, successful treatment or prevention of diabetes depends on a better understanding of how these factors influence the gut immune system, pancreas inflammation and β-cell health (FIG.1).
Nutritional modification of autoimmune diabetes: the link between dietary antigens - gut immune system - islet β-cells
The gut immune system is abnormal in a significant subset of patients with T1D who display one or more of the following: impaired oral tolerance [8], increased gut permeability, and a pro-inflammatory cytokine profile. In the diabetes-prone BB rat, there is a pro-inflammatory Th1 cytokine pattern in the immune cells that infiltrate the islets. This inflammation is dampened in animals fed a protective, antigen-free diet [9] suggesting that diet controls the islet-directed inflammatory process. By contrast, animals fed diabetes-promoting, cereal-based diets display an unusually high proportion of CD4+, IFN-γ producing T cells in the main inductive site for the gut immune system, the mesenteric lymph nodes (MLN) [6]. The findings in this animal model of T1D appear to reflect the situation in humans with T1D. We found that there is a large subset of patients who display strong pro-inflammatory, mainly Th1 cell reactivity to wheat proteins [7]. Immune reactivity to a protein we discovered by screening a wheat cDNA expression library with serum from diabetic BB rats [10], is also increased in a subset of T1D patients [8] and may even be an early marker of celiac disease [11]. Thus, the basis of abnormal immune response to wheat peptide antigens in diabetes-prone animals and humans should be an expanded research focus.
A gut-centric working model of T1D – M2 macrophages and antimicrobial proteins
The gut is where dietary molecules, microbes and other non-self chemicals have their first major encounter with the largest population of immune cells in the body. We hypothesized that the gut barrier and the gut immune system are abnormal in some susceptible individuals permitting uncontrolled amounts of dietary and microbial antigens to stimulate an abnormal immune system to mistakenly target insulin-producing β-cells. This model was described in more detail in two reviews [2,12] and in FIG. 1. Furthermore, our additional studies indicated that there is a deficit of CD163+ M2 immunosuppressive macrophages in the gut of diabetes-prone rats and an immune response against these cells is sometimes observed in humans [13]. Additional studies indicated the involvement of antimicrobial proteins in the development of T1D.
Compromised regenerative capacity and abnormal β-cell metabolism?
Our data showed that diets with different diabetes-promoting capacity affected the ability of the endocrine pancreas to retain sufficient β-cells to maintain normal blood glucose. Two regenerative processes are increased in the pancreas of diabetes-prone rats. The first is islet neogenesis (the formation of β-cells from non-β-cells, e.g. ducts), which is represented by extra-islet, insulin+ clusters. The second is the formation of neogenic structures called tubular complexes [3]. Although both of these processes are increased in the pancreas of diabetes-prone rats, the animals are unable to maintain normal blood glucose levels in the face of a chronic immune attack, possibly because of a decreased capacity to expand into mature islets [5]. These processes represent early but futile attempts to maintain islet mass and could be influenced by diet. Therefore, diets that influence diabetes development probably do so through combined effects on islet β-cell health and the gut immune system. In addition, we reported that aggregates of the normal prion protein, PrPC, occur in the cytosol of β-cells and their formation is responsive to changes in blood glucose [14,15]. This suggested that PrPC can exist in an aggregated form in vivo, and is linked with glucose homeostasis. This is consistent with findings of altered ER stress, metabolism and apoptosis in very young diabetes-prone BBdp rats (16).
Developing a model of T1D pathogenesis that involves early dysregulated metabolism in pancreas, gut immune compartment and other organs. β-cell metabolism could be different in diabetes-prone individuals.
We developed a disease model that integrates the role of gut lumen antigens from diet and microbes, the gut immune system, the endocrine pancreas and the liver in the development of diabetes (16-18). Our research suggested that changes occur very early in the pancreas of diabetes-prone animals long before immune activation occurs. There are indications that other organs such as the gut and liver (18) have diminished capacity to maintain their usual tolerogenic state which leads to beta-cell-specific immune activation by dietary and microbial molecules from the gut lumen. Dr. Scott’s model of type 1 diabetes development proposes that early exposure to various gut lumen antigens has a marked effect on pancreas biology that depends of defects in gut barrier and immune function. These events could be closely linked with other organs (18).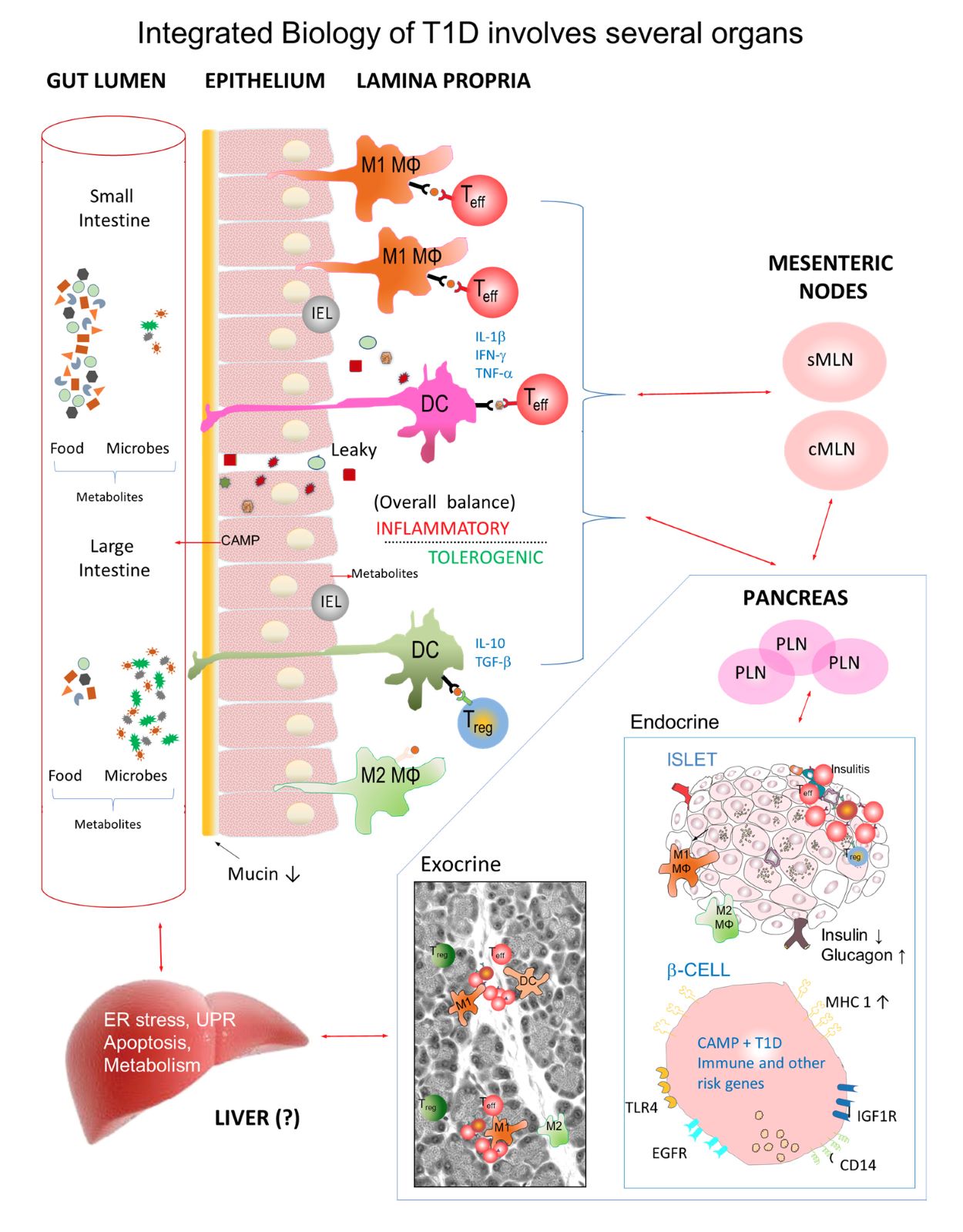
Fig. 1. Role of the gastrointestinal tract, its contents, gut immune system, pancreas and liver in T1D. T1D risk is strongly influenced by environmental factors from the gut lumen that stimulate immune cells to traffic to mesenteric (MLN), pancreatic lymph nodes (PLN) and islets. Diabetes-prone rats fed a protective (antigen-free) hydrolyzed casein (HC) diet have low diabetes incidence compared with animals fed a cereal-based diet. Both diabetes-prone rats and humans display sporadic gut inflammation and leakiness, impaired oral tolerance and low levels of gut CD163+ M2 anti-inflammatory macrophages which can be modified by microbes and dietary antigens. When diabetes is prevented by upregulating the CD163 / heme oxygenase-1 (HO-1) pathway, M2 and M2-like macrophages may be increased and appear in the interstitium, exocrine pancreas and islets. Animals fed a protective HC diet show increased islet-mass compared with cereal-fed rats possibly involving M2 macrophages and antimicrobial factors such as cathelicidin antimicrobial protein (CAMP). Our data suggest there is a prediabetes gene signature in diabetes-prone neonates in both the pancreas and liver (18). Some gene changes occur in the liver before they are seen in the pancreas (see Crookshank et al., 2018). It will be important to consider not only the immune system but also very early changes in pancreas and other organs in T1D pathogenesis (16-18).
- Patrick C, Wang GS, Lefebvre DE, Crookshank JA, Sonier B, Eberhard CE, Mojibian M, Kennedy CR, Brooks SP, Kalmokoff ML, Maglio M, Troncone R, Poussier P, Scott FW: Promotion of autoimmune diabetes by cereal diet in the presence or absence of microbes associated with gut immune activation, regulatory imbalance and altered cathelicidin antimicrobial peptide. Diabetes, 2013
- Lefebvre DE, Powell KL, Strom A, Scott FW: Dietary proteins as environmental modifiers of type 1 diabetes mellitus. Annu Rev Nutr 26:175-202, 2006
- Wang GS, Rosenberg L, Scott FW: Tubular complexes as a source for islet neogenesis in the pancreas of diabetes-prone BB rats. Lab Invest 85:675-688, 2005
- Strom A, Wang GS, Reimer R, Finegood DT, Scott FW: Pronounced cytosolic aggregation of cellular prion protein in pancreatic beta-cells in response to hyperglycemia. Lab Invest 87:139-149, 2007
- Kauri LM, Wang GS, Patrick C, Bareggi M, Hill DJ, Scott FW: Increased islet neogenesis without increased islet mass precedes autoimmune attack in diabetes-prone rats. Lab Invest 87:1240-1251, 2007
- Chakir H, Lefebvre DE, Wang H, Caraher E, Scott FW: Wheat protein-induced proinflammatory T helper 1 bias in mesenteric lymph nodes of young diabetes-prone rats. Diabetologia 48:1576-1584, 2005
- Mojibian M, Chakir H, Lefebvre DE, Crookshank JA, Sonier B, Keely E, Scott FW: A diabetes-specific HLA-DR restricted pro-inflammatory T cell response to wheat polypeptides in tissue transglutaminase antibody negative patients with type 1 diabetes. Diabetes 58:1789-1796, 2009
- Simpson M, Mojibian M, Barriga K, Scott F, Fasano A, Rewers M, Norris J: An exploration of Glo-3A antibody levels in children at increased risk for type 1 diabetes mellitus. Pediatr Diabetes 10:563-572, 2009
- Scott FW, Rowsell P, Wang GS, Burghardt K, Kolb H, Flohe S: Oral exposure to diabetes-promoting food or immunomodulators in neonates alters gut cytokines and diabetes. Diabetes 51:73-78, 2002
- MacFarlane AJ, Burghardt KM, Kelly J, Simell T, Simell O, Altosaar I, Scott FW: A type 1 diabetes-related protein from wheat (Triticum aestivum). cDNA clone of a wheat storage globulin, Glb1, linked to islet damage. J Biol Chem 278:54-63, 2003
- Taplin CE, Mojibian M, Simpson M, Taki I, Liu E, Hoffenberg EJ, Norris JM, Scott FW, Rewers M: Antibodies to the wheat storage globulin Glo-3A in children before and at diagnosis of celiac disease. J Pediatr Gastroenterol Nutr 52:21-25, 2011
- Sonier B, Patrick, C., Ajjikuttira, P. Scott, F.W.: Intestinal Immune Regulation as a Potential Diet-Modifiable Feature of Gut Inflammation and Autoimmunity. International Reviews of Immunology 28:414-445, 2009
- Sonier B, Strom, A., Wang, G.-S., Patrick, C., Crookshank, J.A., Mojibian, M., MacFarlane, A.J., Scott, F.W.: Antibodies from a patient with type 1 diabetes and celiac disease bind to macrophages that express the scavenger receptor CD163. Canadian Journal of Gastroenterology 25:327-329, 2011
- Strom A, Wang GS, Picketts DJ, Reimer R, Stuke AW, Scott FW: Cellular prion protein localizes to the nucleus of endocrine and neuronal cells and interacts with structural chromatin components. Eur J Cell Biol 90:414-419, 2011
- Strom A, Wang GS, Scott FW: Impaired glucose tolerance in mice lacking cellular prion protein. Pancreas 40:229-232, 2011
- Changes in insulin, glucagon and ER stress precede immune activation in type 1 diabetes. Crookshank JA, Serrano D, Wang G-S, Patrick C, Morgan BS, Paré M-F, Scott FW Journal of Endocrinology August, 2018 PMID: 30139929
- Where genes meet environment - Integrating the role of gut luminal contents, immunity and pancreas in type 1 diabetes. Scott FW, Pound LD, Patrick C, Eberhard C, Crookshank JA. Translational Research 179: 183-198, 2017 PMID: 27677687
- Dysregulated liver lipid metabolism and innate immunity associated with hepatic steatosis in neonatal BBdp rats and NOD mice. Serrano D, Crookshank JC, Morgan BS, Mueller RW, Paré M-F, Marandi L, Poussier P, Scott FW Scientific Reports 2019 Oct 10;9(1):14594. doi: 10.1038/s41598-019-51143-7
Selected Publications
Changes in insulin, glucagon and ER stress precede immune activation in type 1 diabetes. Crookshank JA, Serrano D, Wang G-S, Patrick C, Morgan BS, Paré M-F, Scott FW Journal of Endocrinology August, 2018 PMID: 30139929
Where genes meet environment - Integrating the role of gut luminal contents, immunity and pancreas in type 1 diabetes. Scott FW, Pound LD, Patrick C, Eberhard C, Crookshank JA. Translational Research 179: 183-198, 2017 PMID: 27677687
Cathelicidin antimicrobial peptide: a novel regulator of islet function, islet regeneration and selected bacteria Pound LD, Patrick C, Eberhard C, Mottawea W, Wang GS, Abujamel T, Vandenbeek R, Stintzi A, Scott FW Diabetes 64:4135-47, 2015 PMID: 26370175
Jörns A, Arndt T, Vilsendorf AM, Klempnauer J, Wedekind D, Hedrich HJ, Marselli L, Marchetti P, Harada N, Nakaya Y, Wang GS, Scott FW, Gysemans C, Mathieu C, Lenzen S. Islet infiltration, immune cell activation and β-cell death in the NOD mouse, BB rat, Komeda rat, and LEW.1AR1-iddm rat in comparison with the pancreas of patients with type 1 diabetes. Diabetologia, 57:512-21 2014 PMID: 24310561
Patrick C, Wang GS, Lefebvre DE, Crookshank JA, Sonier B, Eberhard C, Mojibian M, Kennedy C, Brooks SPJ, Kalmokoff M, Maglio M, Troncone R, Poussier P, Scott FW. Promotion of autoimmune diabetes by cereal diet in the presence or absence of microbes associated with gut immune activation, regulatory imbalance and altered cathelicidin antimicrobial peptide. Diabetes 62:2036–2047, 2013 PMID: 23349499
Mojibian M, Chakir H, Lefebvre DE, Crookshank JA, Sonier B, Keely E, Scott FW. A diabetes-specific HLA-DR restricted pro-inflammatory T cell response to wheat polypeptides in tissue transglutaminase antibody negative patients with type 1 diabetes. Diabetes 58: 1789-1796, 2009 (Commentary, p. 1723) PMID: 19401421
MacFarlane AJ, Burghardt KM, Kelly J, Simell T, Simell O, Altosaar I, Scott FW A type 1 diabetes-related protein from wheat (Triticum aestivum): cDNA clone of a wheat storage globulin, Glb1, linked to islet damage J. Biol. Chem. 278:54-63, 2003 (Cover Article) PMID: 12409286